
进化论为什么可以解释生物学上的一些“现象”
生命世界的最高分类

俄裔美国果蝇遗传学家 Dobzansky 曾说过:“没有进化,生物学中的所有现象都无法得到合理解释。”我借用了这句话:如果我们不了解特定古老环境中生物的进化史,今天的分子生物学研究将毫无意义。正如达尔文所指出的,我们应该根据生物的自然血统对生物进行分类。如果对生物的分类、识别和命名足够准确,就可以反映生物的进化过程。这是完全有可能的,因为许多关于过去的信息都包含在今天的生物体中。直到最近,人们才意识到微生物学(广义上讲,对细菌及其原生动物后代的研究)对进化的贡献。微生物细胞是生命不可或缺的一部分,因此人们必须越来越关注自身及其对生物进化的重要性。
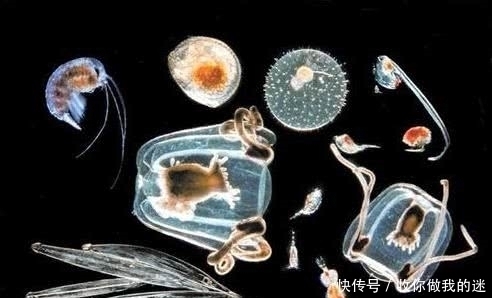
生命世界可以分为两个不同的群体:原核生物和真核生物。尽管卡尔沃斯曾经提出了一个广为接受的基于 16srRNA 的“三域”分类体系,但地球上只有两种基本生命形式。没有化??石记录或生物证据偏离“原核-真核”规则(即没有证据表明这两个种群有原始祖先。原核生物和真核生物的区别是生命世界的最大差距。
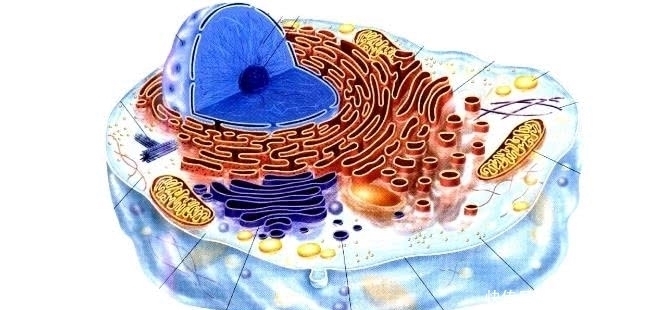
查尔顿是第一个识别并列出古细菌、真细菌和真核生物之间差异的代尔夫特微生物研究所。显然,这两个原核生物群之间的关系比它们与任何真核生物的关系更密切。细菌细胞(其基因组被染色的细丝没有生物膜涂层)或有丝分裂细胞(其蛋白质化的染色体核被脂蛋白生物膜包裹)是所有生命的基本单位。
无论是比活细胞更复杂的物质还是能量流体系统,细胞核是真核生物和细菌的唯一区别。细菌细胞的起源相当于生命本身的起源,而连续内共生理论则通过共生阐明了后续真核生物的起源。
在进一步讨论之前,有必要将生态学中的“共生”概念与进化生物学中的“共生起源”区分开来。
共生是指不同的物种生活在一起,而内共生作为一种拓扑条件,是指一个成员与另一个成员之间形成的共生关系。共生通常是环境融合的结果,但并非总是如此。共生不是进化过程,而是由环境引起的生物体之间的暂时或拓扑关系。共生起源是指长期共生以产生新的组织、器官、生理过程和其他新特征。真核生物的两个主要细胞器,质体和线粒体,是从共生起源进化而来的。
异养需氧蛋白酶可以通过移动厌氧原生动物(例如活的变形鞭毛)而被吞噬。
在选择的影响下,遗传和新陈代谢退化,曾经自由生存的真菌进化成我们今天看到的线粒体。这种共生的产物包括绝大多数异养原生动物,如变形虫、隐生菌、毛滴虫和马铃薯真菌,如疫霉属。同样,一些移动的原生动物以蓝藻细胞为食,但无法成功消化它们,最终形成共生关系。在光线充足的水域中,自然选择再次减少遗传和代谢冗余,并将难以消化的蓝藻与摄入的原生动物永久结合。通过这种方式,产生了在细胞质中含有光合细胞器的藻类。经过进化,一些群体最终成为陆生植物的祖先。鼎福门(包括引起疟疾的疟原虫属在内的一个家族)显然是从这种藻类的谱系进化而来的。该家族的成员,包括弓形虫,保留了一个质粒 DNA 残基,但它们不再具有光合作用的能力。可见“先用后弃”的原则。自然选择没有远见。蓝藻的质体在进化过程中由于无用而逐渐退化。一些自由生活的细菌(如蓝藻)与一些细胞器(如绿藻叶绿体)非常相似,这支持了某些细菌在其他细胞中已经存在数百万年的观点。
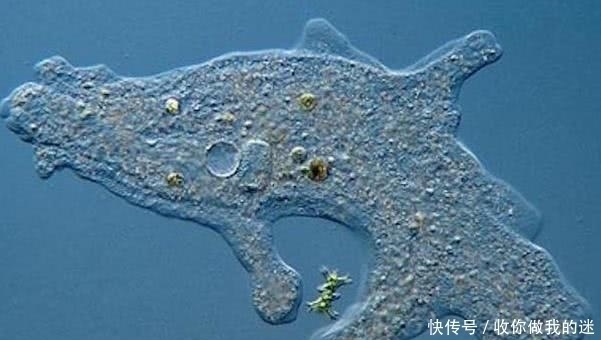
由于线粒体是从自由生活的变形杆菌和自由生活的蓝细菌的质体中获得的,因此没有人质疑有氧呼吸和光合细胞器的共生起源。
我们可以对现代共生关系进行实验分析。内源共生和外源共生对于理解进化过程都很重要。细胞如何融合和减少冗余与第一批真核生物(显然是第一批原生动物)的出现密切相关。具有讽刺意味的是,尽管大多数疾病状况因循环共生而异,但很少有史前生物学家和微生物学家熟悉分析这种几乎无处不在的关系的富有洞察力和蓬勃发展的文献。
在我们对复杂真核生物起源的假设过程中,没有“缺失环节”,每一个假设事件都可以在再生微生物中观察到。基因组学和蛋白质组学研究将证实或证伪这一主要以生物学为基础的进化史。个体生物学和化石记录的直接知识对于重建进化过程至关重要。分子生物学和序列分析不足以建立可验证的进化假设。
下一篇:没有了